MHC paralogon Co-duplicated groups
It was proposed that elucidation of intra-genomic syntenic regions through map self-comparison approach does not provide compelling support for the proposed mechanism of origin of paralogons (Abbasi, 2008). To present a plausible explanation of the evolutionary events that shaped the syntenic relationships seen on present day human MHC bearing chromosomes, topology comparison approach was employed for triplicated/quadruplicated gene families residing on this paralogon (Abbasi, 2010b; Abbasi and Grzeschik, 2007; Zhang and Nei, 1996). Gene families belonging to a particular co-duplicated group suggest that they share similar evolutionary history and might have originated through gene cluster duplication event, whereas genes belonging to different co-duplicated groups may not share their evolutionary history and might not have duplicated simultaneously. Duplication history summary of families residing on MHC paralogon with respect to speciation events and identified co-duplication events are given in the following table.
| Chromosomes/Paralogon | |||||||||
| Name of Co-duplicated group | Co-duplicated Group A | Co-duplicated Group B | Co-duplicated Group C | Co-duplicated Group D | Co-duplicated Group E | Co-duplicated Group F | Remaining Families | Largely duplicated in invertebrates (prior to vertebrate invertebrate split) | Total |
| No. of gene families | 3 | 2 | 13 | 9 | 5 | 2 | 3 | 4 | 40 |
For 40 multigene families residing on MHC bearing paralogon (HSA 1/6/9/19), 6 distinct co-duplicated groups were identified with gene families diversified within the time window of vertebrate-invertebrate and tetrapod-bony fish split. First co-duplicated group presented a topology of type ((Hsa9 Hsa19/Hsa1)(Hsa1/Hsa19 Hsa15/Hsa19)) and suggests the simultaneous duplication of 3 gene families (Figure 1-A). Second co-duplicated group comprised members from 2 gene families and shows a topology of type ((Hsa6/Hsa3 Hsa1)Hsa1/Hsa7)(Figure 1-B). Third co-duplicated group was the largest with 13 gene families and shows a topology of type (((Hsa9 Hsa19/Hsa6)Hsa1/Hsa19)Hsa1/Hsa15)(Figure 1-C). Fourth co-duplicated group included 9 gene families and follows a topology of type ((Hsa1/Hsa15 Hsa19)Hsa9/Hsa6)(Figure 1-D). Fifth co-duplicated group shows a topology of type ((Hsa1/Hsa19 Hsa9)Hsa6/Hsa19) and involved gene members from 5 families (Figure 1-E).Sixth co-duplicated group comprised members from 2 gene families and shows a topology of type ((((Hsa6/Hsa1 Hsa9/Hsa6)Hsa19/Hsa9)Hsa1/Hsa11)Hsa19)(Figure 1-F).(Naz et al, 2017).
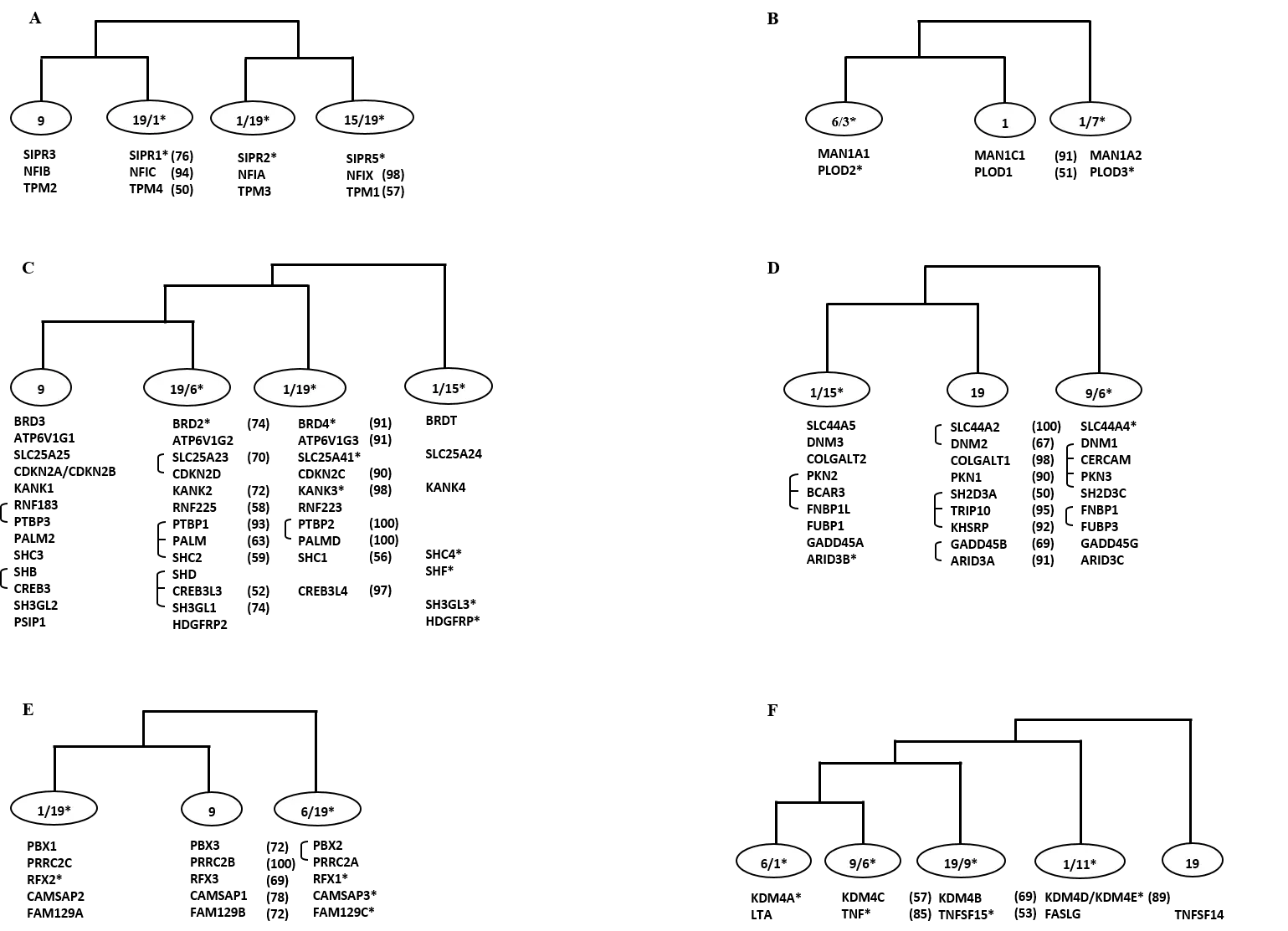
Figure 1: Schematic topologies indicating consistencies in phylogenies of families including members on at least three of the human 1, 6, 9, 19 chromosomes. In each case the percentage bootstrap support is given in parentheses and only values ≥50% are presented here. The connecting bars on the left depict the close physical linkage. * Represents situation where a gene family member is not residing on mentioned chromosomes.
Useful references: